

 当前位置:首页 > 新闻中心 > 新闻资讯
当前位置:首页 > 新闻中心 > 新闻资讯
非酒精性脂肪性肝病(NAFLD)对公众健康构成严重威胁,其潜在机制尚不清楚。先前的研究利用基因重要性计算器(GIC)筛选重要基因时,预测核糖体修饰蛋白rimk样家族成员A (RIMKLA)是一个重要基因,但其功能仍不清楚。
2024年8月8日,北大基础医学院杨吉春教授、北大人民医院迟毓婧副教授联合吉林大学王国庆教授、吉林大学第一医院刘兴凯教授团队在Signal Transduction and Targeted Therapy(IF 40.8)上在线发表了题为“Ribosomal modification protein rimK-like family member A activates betaine-homocysteine S-methyltransferase 1 to ameliorate hepatic steatosis”的文章,研究发现RIMKLA是一种新型蛋白激酶,可使BHMT1 Thr45位点磷酸化,从而抑制脂质合成和摄取。在肥胖情况下,抑制RIMKLA会损害BHMT1活性,从而促进肝脏脂质沉积,表明激活肝脏RIMKLA-BHMT1功能复合物可能是治疗高同型半胱氨酸血症和代谢紊乱的新策略。


01
1、肝脏RIMKLA过表达可改善肥胖小鼠的高血糖和脂肪变性
研究人员使用之前研发的基因重要性计算器(GIC)筛选重要基因时,核糖体修饰蛋白rimk样家族成员A (RIMKLA)被预测为一个必需基因,进一步检测发现RIMKLA在患有NAFLD的小鼠和人肝脏中表达降低,数据显示RIMKLA可能在调节肝脏糖脂代谢中发挥作用。为了评估肝脏RIMKLA对糖脂代谢的作用,研究人员利用Ad-RIMKLA对HFD喂养3个月的小鼠进行RIMKLA过表达处理。与对照组小鼠相比,RIMKLA过表达小鼠表现出空腹血糖水平、葡萄糖耐受不良、肝脏葡萄糖生成和胰岛素抵抗的显著改善;RIMKLA过表达小鼠肝脏细胞质和细胞核中RIMKLA的表达均增加,肝脏脂质积累减轻,肝脏甘油三酯(TG)含量显著降低,血清TG水平略有升高。此外,肝脏RIMKLA过表达降低了血清中谷丙转氨酶( ALT )和谷草转氨酶( AST )的活性。进一步在HFD喂养6个月的小鼠中证实了RIMKLA过表达的作用。考虑到腺病毒转导在体内的过表达时间相对较短,使用AAV8在db/db小鼠肝脏过表达RIMKLA (AAV8-RIMKLA)研究其对糖/脂代谢的长期影响。注射AAV8-RIMKLA 后10周,db/db小鼠胰岛素敏感性、葡萄糖耐受不良、肝糖生成以及空腹高血糖均有显著改善。AAV8-RIMKLA显著降低了db/db小鼠的肝脏脂肪含量和总脂肪体积,肝脏和血清TG含量降低。总体而言,急性或慢性肝脏RIMKLA过表达显著纠正了肥胖小鼠糖脂代谢失调。
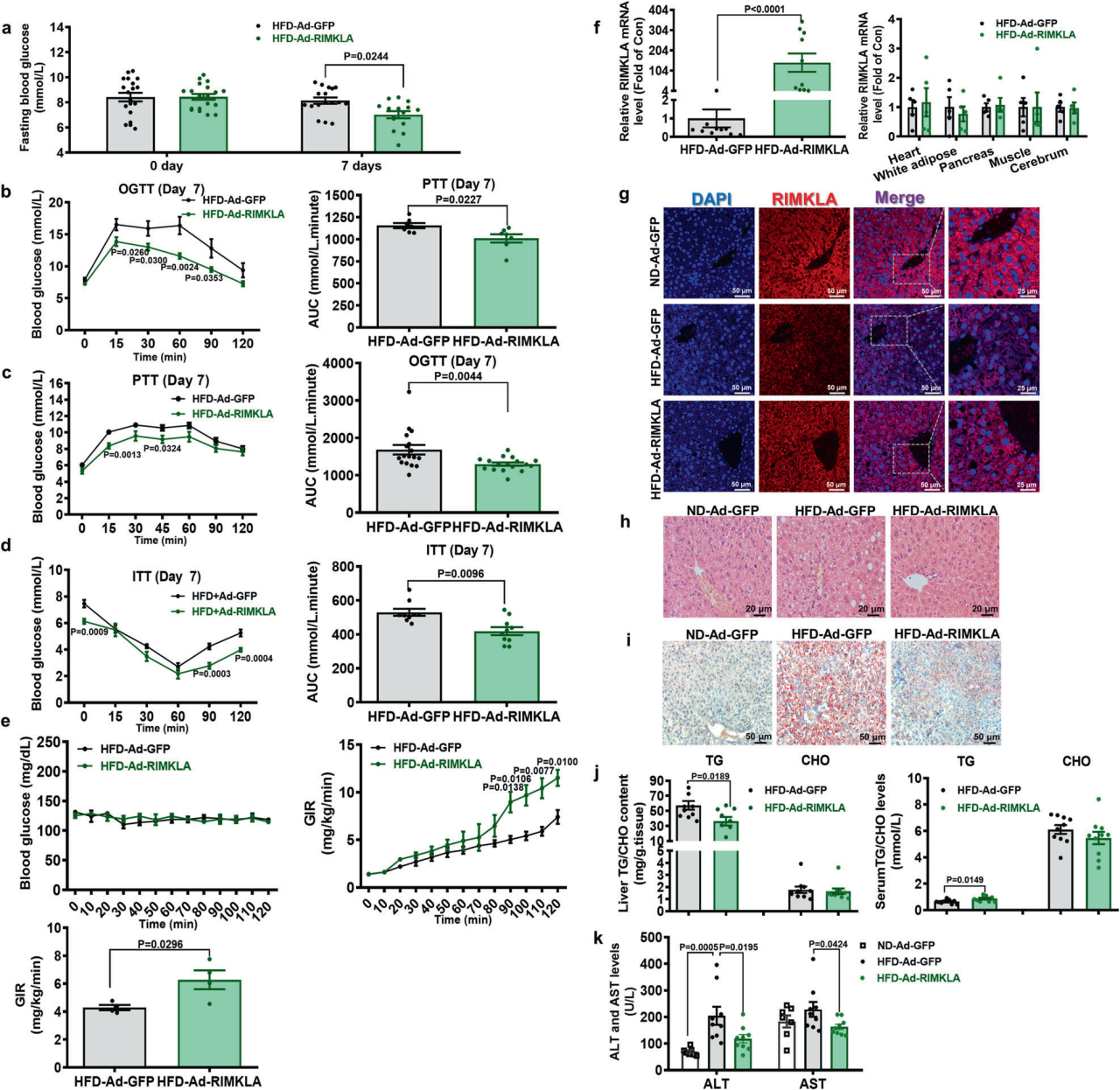
图1. 肝脏RIMKLA过表达可改善HFD小鼠糖脂代谢失调
2、RIMKLA抑制小鼠肝脏和培养肝细胞中的糖异生和脂质生成/脂质摄取基因表达
在糖尿病小鼠肝脏中,注射Ad-或AAV8-RIMKLA可使RIMKLA蛋白水平升高约2倍。RIMKLA过表达增加小鼠肝脏中蛋白激酶B (Akt)磷酸化,降低葡萄糖6磷酸酶(G6Pase) mRNA和蛋白水平。在小鼠肝细胞和HepG2细胞中,RIMKLA过表达增加了磷酸化Akt ( pAkt )的核分布,促进了叉头框蛋白O1 ( FOXO1 )的核排斥,并降低了G6Pase的表达。为了研究RIMKLA在脂质代谢中的作用机制,研究人员评估了RIMKLA过表达对脂质代谢关键基因mRNA水平的影响。在RIMKLA过表达后,HFD小鼠肝脏和FFA诱导的肝细胞中,FASn(关键脂肪生成基因之一)和CD36(关键脂肪酸转运蛋白之一)的mRNA水平均持续降低;同时,RIMKLA过表达降低了肥胖小鼠肝脏、人HepG2细胞和小鼠肝细胞中的FASn和CD36蛋白水平。此外,在FFAs存在的情况下,RIMKLA过表达也降低了小鼠肝细胞中FASn和CD36蛋白水平,表明RIMKLA过表达减少了FFA促进的脂质沉积。以上数据表明,RIMKLA过表达通过抑制脂质从头合成和脂质摄取改善肝脏脂质沉积,并通过抑制FOXO1抑制糖异生。
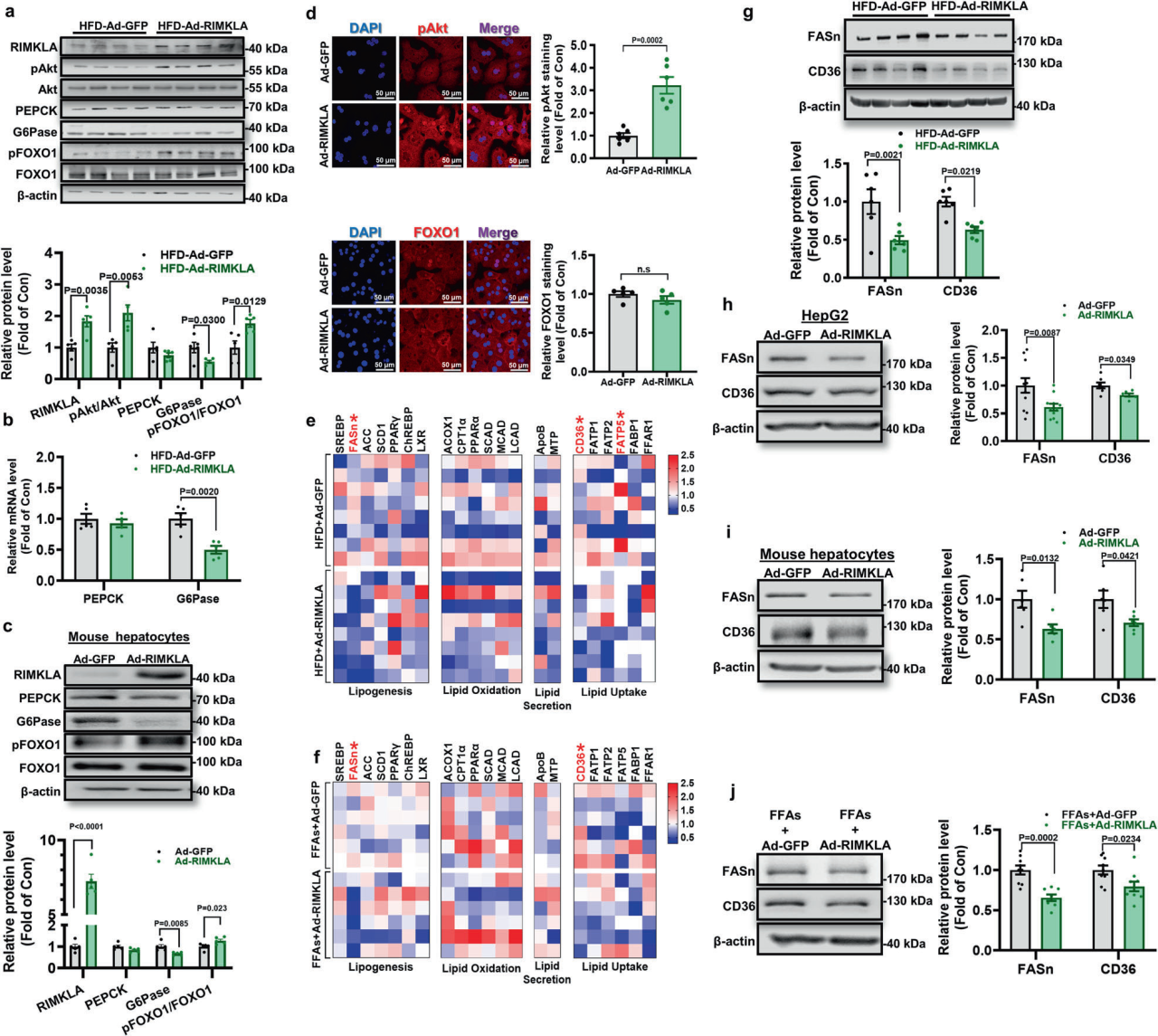
图2. RIMKLA过表达抑制糖异生和脂肪生成/脂质摄取基因的表达
3、RIMKLA通过磷酸化Thr45与BHMT1相互作用并激活BHMT1
为了进一步明确RIMKLA抑制FASn和CD36表达的机制,研究人员在小鼠肝细胞中过表达含有6×His-tag的RIMKLA,然后使用IgG或抗His抗体进行免疫共沉淀(Co-IP),检测到BHMT1可能与其相互作用。已有研究报道BHMT1在调节脂质代谢中发挥重要作用,因此进一步探讨了RIMKLA是否与BHMT1相互作用以调节脂质代谢,并通过实验证实了RIMKLA与BHMT1的相互作用。RIMKLA过表达对小鼠和人肝细胞BHMT1 mRNA和蛋白水平影响不大,但增加了小鼠和人肝细胞中BHMT1的Hcy-Met转化活性,推测RIMKLA可能通过诱导蛋白修饰来激活BHMT1。利用不同生信预测方法发现苏氨酸(Thr) 45位点和丝氨酸(Ser) 79位点是BHMT1蛋白中常见的修饰位点,通过实验证实了RIMKLA直接磷酸化BHMT1的Thr45位点。进一步研究其作用机制,发现RIMKLA通过磷酸化BHMT1的Thr45位点以清除Hcy,并抑制Hcy诱导的AP1激活和FASn、CD36的上调,减少脂质沉积。
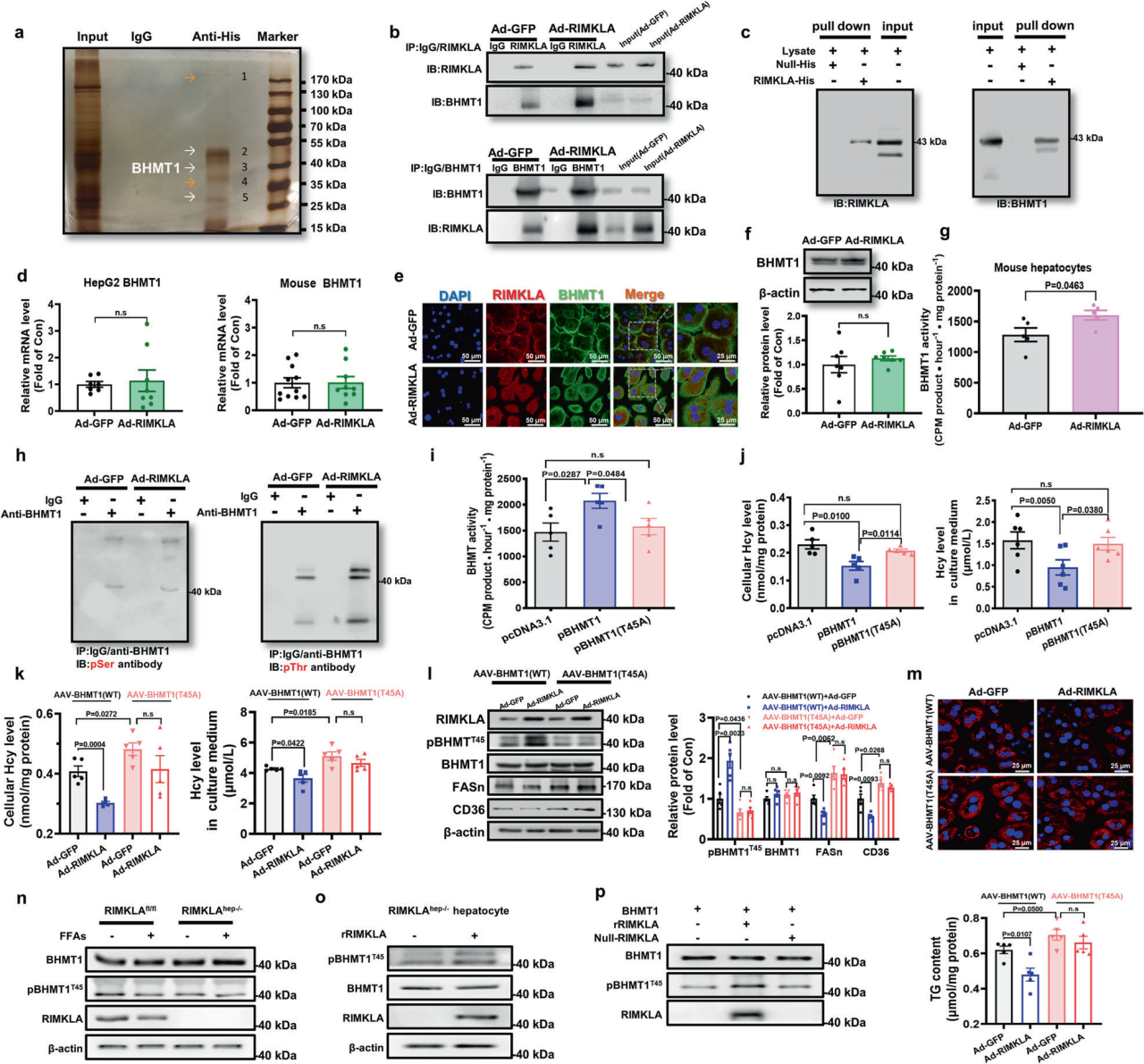
图3.RIMKLA通过磷酸化肝细胞中的Thr45位点与BHMT1相互作用并激活BHMT1
4、肝细胞特异性的RIMKLA缺失会加重HFD小鼠的HHcy、高血糖和脂肪变性
为了进一步证实RIMKLA在调节Hcy和糖/脂代谢中的作用,研究人员构建了肝细胞特异性敲除RIMKLA的小鼠(RIMKLAhep-/-)。HFD喂养8-12周后,RIMKLAhep-/-小鼠表现出比对照组小鼠更严重的葡萄糖耐受不良、肝糖生成和胰岛素抵抗,脂肪含量和总脂肪体积更高。此外,RIMKLAhep-/-小鼠的脂质摄取率高于对照组小鼠。肝细胞中RIMKLA的缺失导致血清和肝脏中Hcy水平升高,Met水平降低;在肝脏和血清CHO水平不变的情况下,RIMKLAhep-/-小鼠的肝脏脂质沉积更为严重。RIMKLAhep-/-小鼠肝脏中BHMT1的磷酸化和酶活性降低,而其蛋白水平保持不变,磷酸化的AP1、FASn和CD36表达增加。与对照小鼠肝细胞相比,培养的RIMKLAhep-/-小鼠肝细胞中Hcy水平、AP1磷酸化、FASn和CD36表达以及脂质沉积增加,但被BHMT1过表达所拯救。以上表明肝细胞特异性RIMKLA缺失会加重HFD小鼠的HHcy、高血糖和脂肪变性。

图4. 肝脏特异性敲除RIMKLA导致代谢紊乱加重
02
综上所述,RIMKLA是一种新型蛋白激酶,能够磷酸化BHMT1的Thr45位点,从而激活BHMT1。在肥胖状态下,高脂血症和高胰岛素血症抑制RIMKLA,使BHMT1活性降低,Hcy水平升高,进而激活AP1,诱导FASn和CD36的转录,刺激脂质从头合成和摄取。RIMKLA-BHMT1轴受损促进脂质和Hcy代谢紊乱之间的恶性循环,从而触发NAFLD和糖尿病的发生和发展。本研究表明激活肝脏RIMKLA-BHMT1功能复合物可能是治疗HHcy和代谢紊乱的新策略。

400-077-2566

service@wzbio.cn








