

 当前位置:首页 > 研究领域 > 神经系统
当前位置:首页 > 研究领域 > 神经系统

应激反应是各种紧张性刺激物引起的个体非特异性反应,包括生理反应和心理反应两大类。生理反应表现为交感神经兴奋、垂体和肾上腺皮质激素分泌增多、血糖升高、血压上升、心率加快等;心理反应包括情绪反应与自我防御反应等。研究发现应激反应会引起外周T淋巴细胞功能紊乱,脑神经可塑性的变化,增加抑郁和焦虑的风险。T淋巴细胞在情感障碍疾患中的调节机制和病理性作用是怎样的呢?浙江大学靳津和东南大学柴人杰研究团队使用维真生物AAV-shRNA产品在《Cell》(IF=38.637)杂志联合发表“Stress-Induced Metabolic Disorder in Peripheral CD4+T Cells Leads to Anxiety-like Behavior”的研究论文,文章揭示了CD4+ T细胞嘌呤代谢紊乱与应激诱导的焦虑行为之间的关联。
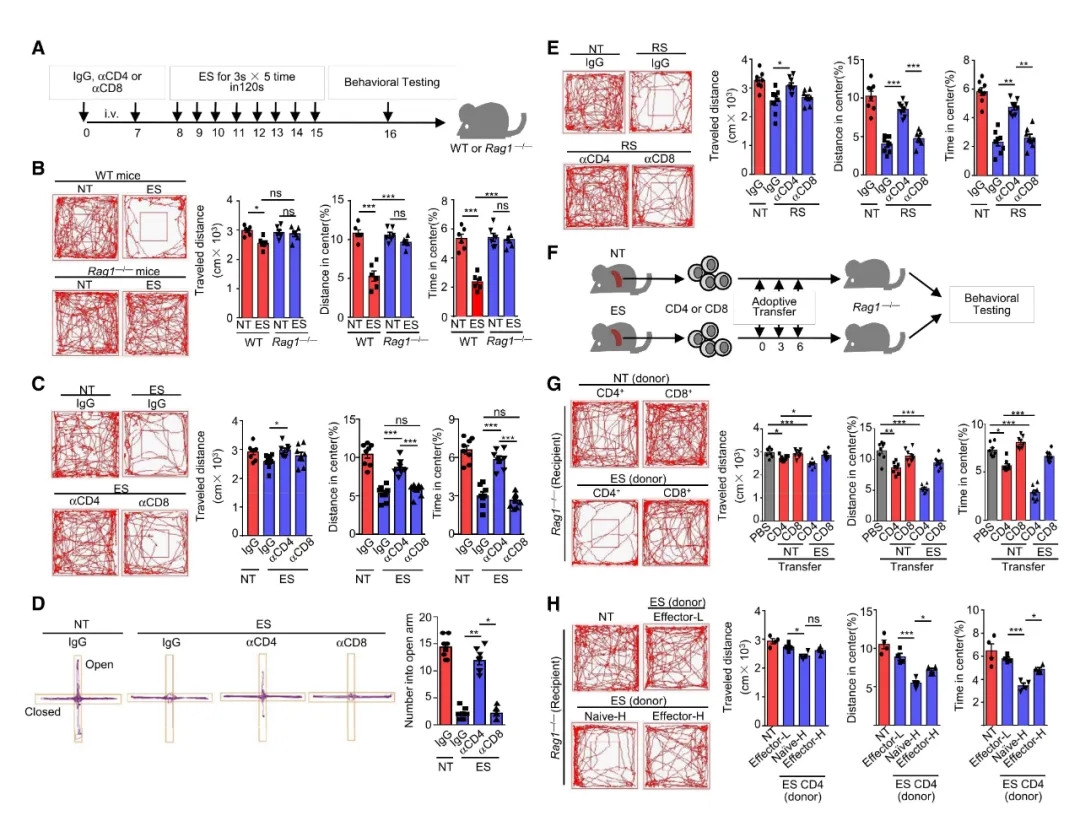
作者通过足部电击 (electronic foot shock,ES)野生型(WT)和Rag1-/- (免疫缺陷)小鼠诱发焦虑模型,开场试验(Open Field Test,OFT)发现,Rag1-/-小鼠对中心区域探索和运动的兴趣没有降低,与研究报道急性ES小鼠的外周血管CD4+和CD8+淋巴细胞数量明显增加结果一致,表明焦虑发生的起始阶段免疫系统会适应性的改变。
通过静脉注射中和抗体来耗尽CD4+和CD8+细胞,作者发现只有CD4+ T细胞的去除显著逆转了OFT和高架十字迷宫实验(elevated plus-maze test,EPM)ES诱导的焦虑行为。CD4+ T细胞缺乏可以阻止急性束缚应激(acute restraint stress,RS)引起小鼠产生焦虑。上述结果表明CD4+ T细胞对物理应激诱发的焦虑行为有着广泛的影响。
为评价T细胞是否具有焦虑印迹保留作用,将NT小鼠和ES小鼠脾脏CD4+或CD8+ T细胞分别过继转移到Rag1-/- 小鼠,发现只有接受ES诱导的CD4+T细胞的Rag1-/-小鼠在OFT中产生焦虑行为,且初始CD4+ T细胞引发的焦虑程度要比效应T细胞更严重。(实验结果如图1)
测序分析ES小鼠的CD4+和CD8+ T细胞RNA转录组,CD4+ T细胞中鉴定出128个特异性差异表达基因(DEGs),基因本体分析显示大多数DEGs编码线粒体蛋白质。胞外酸化速率(extracellular acidification rate, ECAR)和耗氧速率(axygen consumption rate, OCR)测定显示,ES和RS小鼠初始CD4+ T细胞的糖酵解和氧化磷酸化水平均显著下降,线粒体的结构和功能受到影响。通过共聚焦显微成像和免疫印迹技术检测,发现ES初始CD4+ T细胞的线粒体呈点状,线粒体融合的外膜蛋白MFN2和MIGA2显著减少,表明CD4+ T细胞在应激状态下表现为线粒体形态异常和代谢功能障碍。
研究发现白三烯B4(LTB4)给药会使小鼠产生严重的焦虑行为,去除CD4+ T细胞后焦虑行为消失。体外实验显示在应激反应中LTB4显著增加线粒体分裂,降低了MFN2和MIGA2在初始CD4 + T细胞中的表达。研究证明应激反应可诱导LTB4启动外周血CD4+ T细胞线粒体的裂变,并引发焦虑。(实验结果如图2)

为了确定T细胞线粒体形态与焦虑行为的关系,作者建立了Miga2-/-小鼠,OFT、EPM、暗-光转换和悬尾实验中,Miga2-/-小鼠表现出严重的抑郁症状,并在初始CD4+ T中观察到了高度破碎的线粒体。为了进一步阐明外周血CD4+ T细胞线粒体分裂的特殊功能,作者生成了Miga2 T细胞条件敲除(KO) (Miga2TKO)小鼠。缺乏Miga2的T细胞在T淋巴细胞的发育和稳态方面没有表现出任何明显的异常,但Miga2TKO小鼠仍然表现出类似于Miga2-/-小鼠的焦虑样行为。Miga2TKO小鼠在连续EPM测试中表现出正常的恐惧记忆消失,说明Miga2缺陷的CD4+ T细胞对正常的学习记忆没有影响。为了消除Miga蛋白在焦虑中的特殊功能,作者又生成了Mfn1和Mfn2 T细胞条件下双KO (Mfn1/ 2TKO)小鼠,T细胞中缺乏Mfn1/2的小鼠与野生型小鼠相比也表现出类似焦虑的行为,这表明焦虑行为是由线粒体形态紊乱引起的,而不是由某些线粒体蛋白的特定功能引起的。(实验结果如图3)

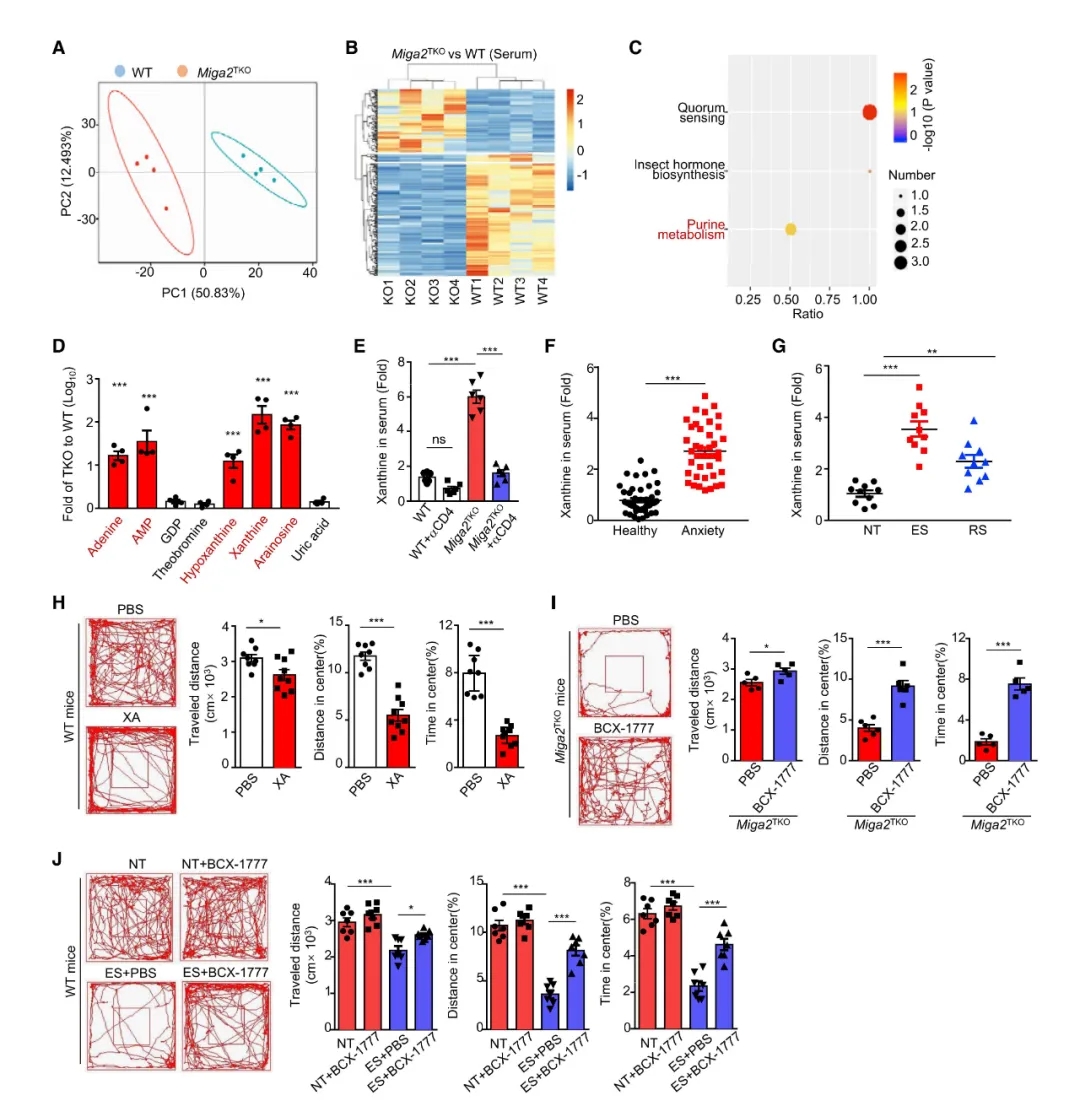
聚类分析显示Miga2TKO小鼠的代谢组学特征与同窝WT小鼠有显著差异,代谢产物的差异主要集中于嘌呤代谢。Miga2TKO小鼠体内嘌呤及其衍生物(腺嘌呤、次黄嘌呤和黄嘌呤)的含量是同窝野生型小鼠的10-100倍。去除CD4+ T细胞后,Miga2TKO小鼠血清黄嘌呤浓度显著降低。在两种啮齿动物焦虑模型和过继性转移ES小鼠CD4+ T细胞的Rag1-/-小鼠血清中也观察到黄嘌呤浓度明细增加。
WT小鼠体内注射合成的黄嘌呤或腺苷发现,黄嘌呤、腺嘌呤和单磷酸阿糖腺苷都能引发小鼠产生焦虑行为。BCX-1777(嘌呤核苷磷酸化酶抑制剂)明显降低Miga2TKO和ES小鼠的焦虑症状,同时显著降低小鼠血清黄嘌呤的浓度。由此可知,病理性CD4+ T细胞引起的黄嘌呤过量在焦虑行为中起至关重要的作用。(实验结果如图4)
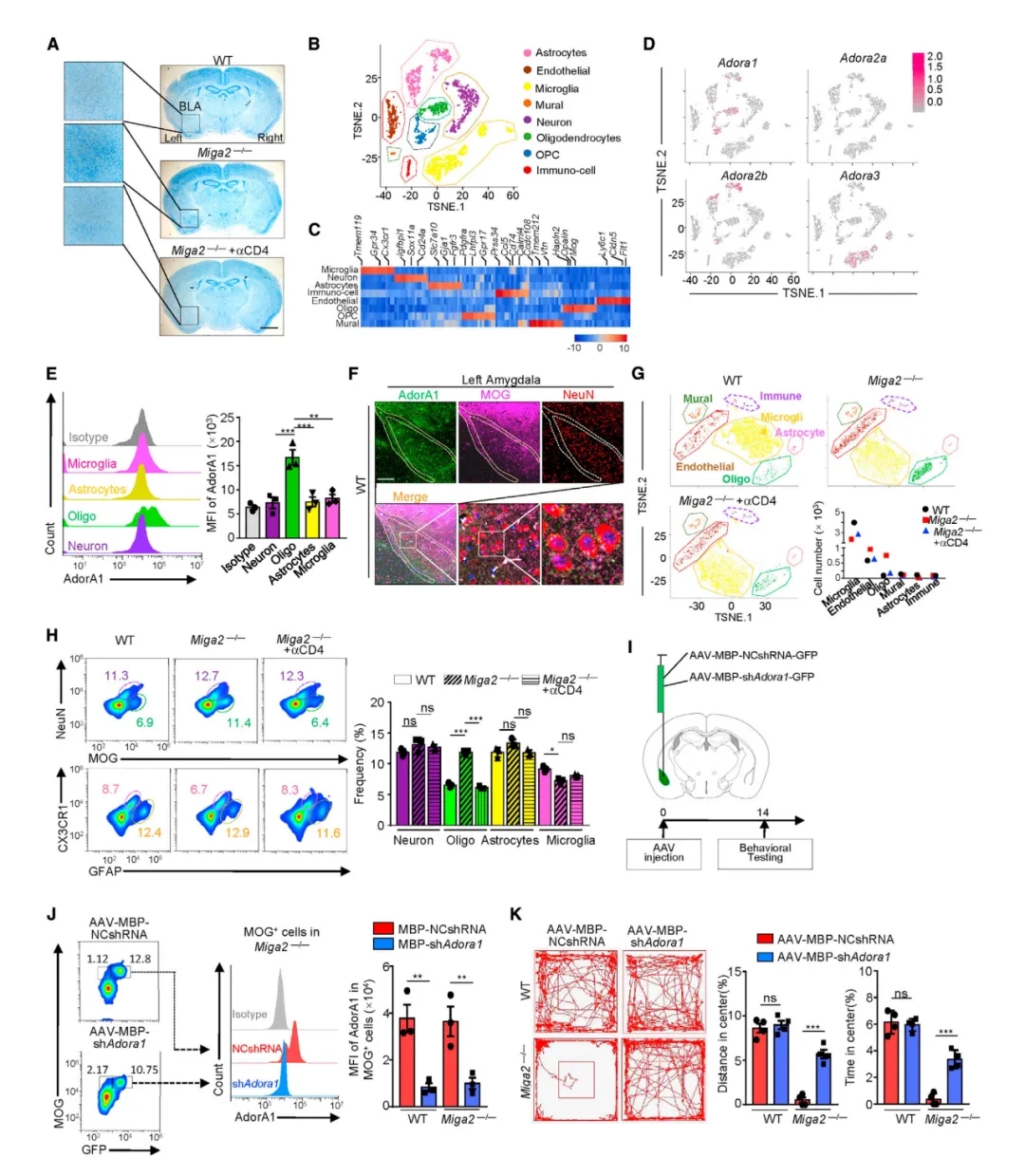
研究表明杏仁核在恐惧和持续焦虑的产生起关键作用,左杏仁核与社交焦虑、强迫症、创伤后应激以及焦虑有关。组织学分析发现,Miga2TKO小鼠左杏仁核比右杏仁核和WT小鼠杏仁核更大,非神经元细胞的数量更多。经黄嘌呤处理的小鼠左杏仁核表现出与Miga2TKO小鼠相似的病理表型。单细胞RNA测序显示,腺嘌呤和黄嘌呤发挥通过A1、A2A、A2B和A3四种受体亚型发挥各自的生理功能,且A1在少突胶质细胞中大量表达。
作者分析了 WT、Miga2-/-、αCD4处理的Miga2-/-小鼠杏仁核中非神经元细胞的转录组。scRNA 序列和FACS分析显示Miga2-/-小鼠少突胶质细胞比例显著增加,去除CD4+ T细胞可逆转此现象。BRdU掺入试验发现,黄嘌呤作为致病因素,可显著增加少突胶质细胞的DNA合成和细胞周期,直接诱发少突胶质细胞的增殖。已有资料表明黄嘌呤作用于A1受体并促进蛋白激酶A (PKA)/ cAMP的活性。作者发现Miga2-/-小鼠少突胶质细胞中PKA/ cAMP通路被明显激活,耗尽CD4+T细胞可下调该通路。作为左侧杏仁核少突胶质细胞唯一可检测到的腺苷受体A1,通过注射腺相关病毒AAV-MBP-shAdor1-GFP(维真生物提供)成功地特异性敲低A1受体,Miga2-/-小鼠不再表现出焦虑症状。(实验结果如图5)
嘌呤有两种合成途径:从头合成和回收。在从头合成途径中,葡萄糖代谢产物5-磷酸糖苷基-1-焦磷酸(PRPP)提供了形成嘌呤环的骨架结构。研究发现Miga2-/-和Mfn1/2TKO小鼠CD4+ T细胞氧化磷酸化和糖酵解活性显著降低,但5-磷酸核糖(R-5-P)、CAIR、腺苷和肌苷则大量增加,表明线粒体在从融合到裂变过程中,葡萄糖通过磷酸戊糖途径(PPP)重新合成嘌呤。2-脱氧-D-葡萄糖(抑制糖酵解、PPP和嘌呤的合成)可使Miga2TKO小鼠的焦虑症状和左杏仁核病理特征恢复正常。
WT和Miga2-/-小鼠初始CD4+ T细胞的转录组分析表明,Miga2-/-小鼠的 CD4 + T细胞与糖酵解和脂肪酸ß氧化途径相关的多个关键酶转录水平显著降低,但是嘌呤合成所需的分子:己糖激酶3 (Hk3)、腺苷脱氨酶(Ada)、嘌呤核苷磷酸化酶2 (Pnp2)和黄嘌呤氧化酶/黄嘌呤脱氢酶(Xdh)有所增加。PNP催化肌苷和鸟苷转化为次黄嘌呤或鸟嘌呤。与PNP相似,PNP2也是一种调节嘌呤代谢途径和黄嘌呤产生的酶,由于其在初始CD4+ T细胞中的低表达,认为其缺乏可能不会像PNP缺乏那样导致严重的免疫缺陷。Pnp2-/-小鼠未表现出任何明显的T淋巴细胞发育或成熟障碍,进一步与Miga2-/-小鼠杂交产生Pnp2-/-/Miga2-/-小鼠,当Pnp2-/-/Miga2-/-CD4+ T细胞过继转入Rag1-/-小鼠后,Pnp2-/-/Miga2-/- CD4+ T细胞不像Miga2-/- CD4+ T细胞那样引起焦虑症状。由于Pnp2缺失不能完全阻断黄嘌呤的合成,在移植Pnp2-/-/Miga2-/- CD4+ T细胞的Rag1-/- 小鼠血清中,黄嘌呤仍有适度升高。这些结果表明CD4+ T细胞来源的过量黄嘌呤可直接导致焦虑行为。(实验结果如图6)

干扰素调节因子-1(IRF-1)作为一种转录因子,参与细胞增殖、分化、凋亡和免疫调节等多种细胞过程。作者发现Miga2-/-和ES小鼠CD4+T细胞中IRF-1显著积累。染色质免疫共沉淀技术研究表明巨噬细胞IRF-1富集于Ada、Xdh和Pnp2的启动子区域。IRF-1缺乏使Miga2-/-小鼠CD4+ T细胞Ada和Xdh mRNA和蛋白水平恢复正常化,减轻了Miga2TKO小鼠的焦虑行为。CD4+ T细胞中的IRF-1在线粒体裂变调节嘌呤合成并产生焦虑症状中起着至关重要的作用。(实验结果如图7)

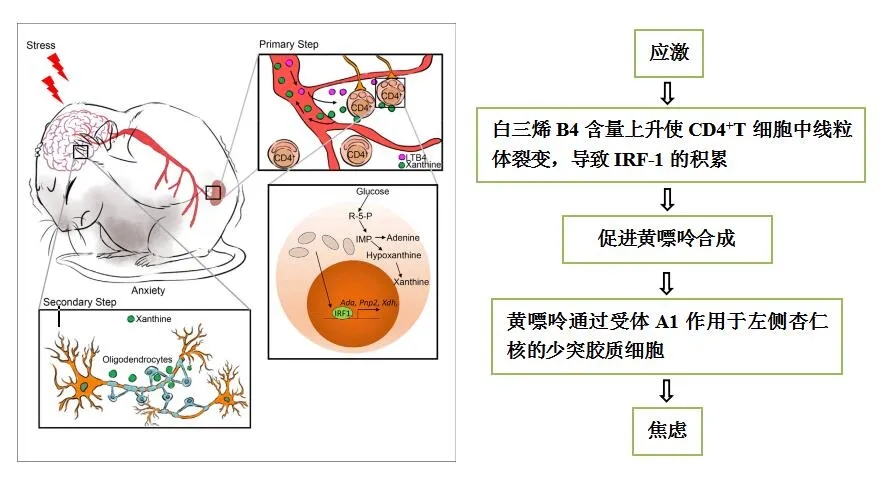
本研究揭示了外周血CD4+ T细胞来源的黄嘌呤和焦虑行为之间的重要联系。如下:
小V相信AAV-shRNA相关产品将对各种精神和代谢疾病药物的研发提供巨大帮助。
靳津教授 现担任浙江大学生命科学研究院教授,长期从事非经典NF-kB的调控机理,以及蛋白修饰在自主免疫性疾病发病过程中调控功能的研究,已在Nature等国际一流期刊发表研究论文30余篇。
柴人杰教授 现任东南大学生命科学研究院教授,近5年发表SCI论文72篇(总影响因子481.8);长期致力于神经元和内耳毛细胞的再生和保护以及神经干细胞和内耳干细胞的转录调控机制的研究。
| AAV9-GFAP-GFP-miR30-shRNA(mAdora1) |
| AAV8-MBP-GFP-miR30-shRNA(mAdora1) |
| AAV8-MBP-GFP-miR30-shRNA(scramble) |
① 现有涵盖27000余种人源基因的siRNAs序列;
② 可随时将shRNAs克隆至腺病毒、慢病毒、腺相关病毒载体,制备病毒载体;
③ 此外,维真还可为您提供专属的shRNA克隆服务和基因沉默效果检测服务。
具体服务内容如下:
shRNA克隆服务:
| 服务编号 | 服务类型 |
|---|---|
| WZ030001 | 单shRNA质粒构建 |
| WZ030002 | 套餐shRNA质粒构建(3保1) |
| WZ030003 | 套餐shRNA质粒构建(4保1) |
| WZ030004 | 2 in 1 shRNA质粒构建 |
| WZ030005 | 3 in 1 shRNA质粒构建 |
| WZ030006 | 4 in 1 shRNA质粒构建 |
| WZ030007 | 诱导性shRNA质粒构建(Dox、Cre……) |
| WZ030008 | mir30-based shRNA质粒构建 |
基因沉默效果检测服务:
| 服务编号 | 服务类型 | 检测方法 |
|---|---|---|
| WZ090001 | 特异shRNA沉默效果检测 | 实时定量PCR推荐!Western blot |
| WZ090002 | 高效shRNA筛选 | 实时定量PCR推荐!Western blot |
欲知更多服务详情请致电400-077-2566。

400-077-2566

service@wzbio.cn







