

 当前位置:首页 > 研究领域 > 神经系统
当前位置:首页 > 研究领域 > 神经系统
睡眠是生命所需。人一生中有将近1/3的时间处于睡眠状态。睡眠质量的好坏与人体健康密切相关。睡眠障碍会引起负面情绪,严重时会引发重大精神疾病。但是人为什么需要睡眠?熬夜后为什么睡得更沉、更香?睡眠到底是如何被调控的呢?一系列科学问题尚待解决。
人体内有两大系统会引发睡意:睡眠稳态与昼夜节律。昼夜节律是通过内在的生物钟控制一天中睡眠觉醒的时间;睡眠稳态则主要是由睡眠压力进行调控,睡眠压力的累计程度控制机体的睡眠量。先前研究已鉴定了许多睡眠稳态因子和生物学过程。其中,基底前脑(basal forebrain,BF)腺苷的释放是睡眠稳态的重要生理调节因子。脑内与睡眠觉醒相关的神经核团有很多,BF是调控该周期的一个重要核团。BF内含有胆碱能、谷氨酸能和GABA能多种类型神经元,这些神经元活动如何调控腺苷的释放仍是一个谜。
近日,北大李毓龙课题组与中科院脑智卓越创新中心徐敏课题组合作在《Science》发表了题为《Regulation of sleep homeostasis mediator adenosine by basal forebrain glutamatergic neurons》的研究论文。该项研究利用新型遗传编码的高灵敏性、高特异性和高时间分辨率的腺苷探针,联合运用光纤记录、光遗传学和细胞特异损毁技术揭示了BF谷氨酸能神经元在调控腺苷动态变化和睡眠稳态中的关键作用,进一步阐明了睡眠稳态调控的神经环路机制,为探索睡眠障碍的治疗方法提供了重要参考。
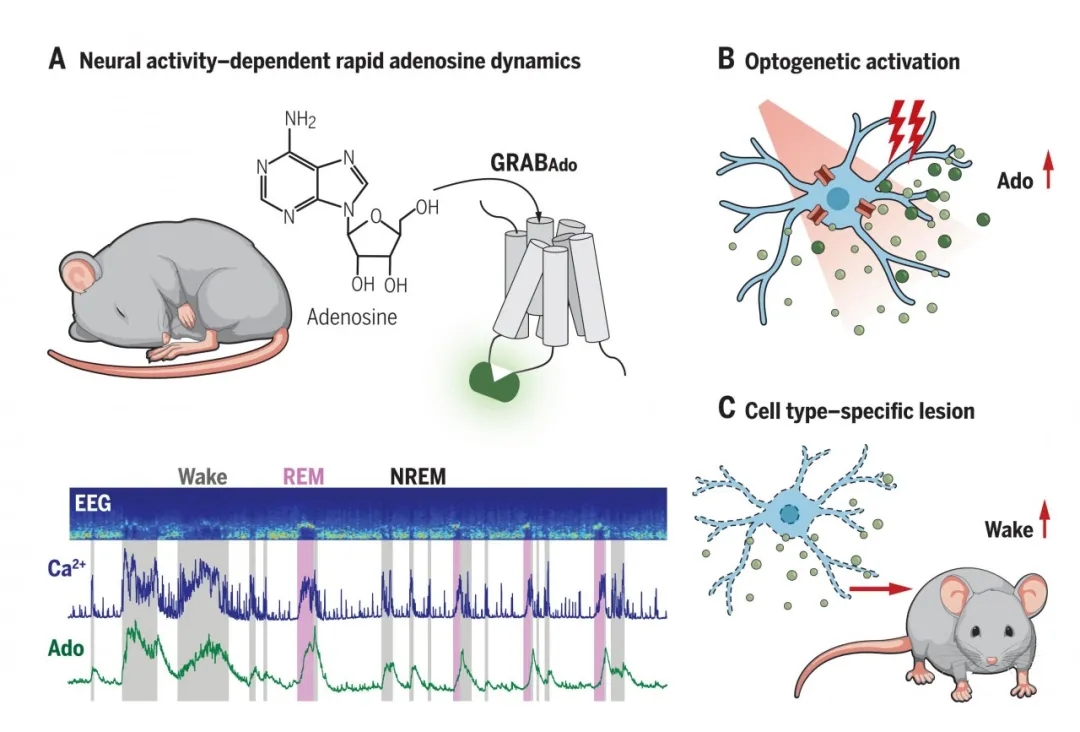
为了高特异性和高时间分辨率检测睡眠觉醒周期中BF胞外腺苷浓度的动态变化,李毓龙教授组开发了腺苷探针(GRABAdo),该探针的检测原理是基于腺苷G蛋白偶联受体的激活,使胞外腺苷的浓度通过绿色荧光蛋白的荧光强度来测量。作者从灵敏性、特异性和胞内信号通路的偶联等方面分别进行了实验,发现迭代优化后的GRABAdo1.0(简称Ado1.0)具有较高的灵敏性和特异性,同时与胞内信号通路具有微乎其微的偶联,对细胞生理无明显影响(图1)。

据睡眠中脑电图表现、眼球运动情况和肌张力变化,睡眠可分为非快速眼球运动睡眠 (NREMS)和快速眼球运动睡眠(REMS)两种时相。其中,REMS由于持续时间短,传统的微透析无法记录腺苷浓度的变化。腺苷探针的高时间分辨率使问题得到了有效解决。作者使用AAV2/9-hsyn-GRABAdo1.0-mScarlet和AAV2/9-hsyn-GRABAdo-mut脑立体定位注射小鼠BF并测量荧光信号。为了排除运动噪音,作者将Ado1.0与红色荧光蛋白mScarlet融合表达,该红色荧光蛋白对腺苷浓度的变化不敏感。实验结果显示:小鼠BF胞外腺苷浓度的变化顺序为:快速眼动睡眠阶段>觉醒阶段>非快速眼动睡眠阶段(后2个阶段的结果跟传统微透析的结果一致)。而且,当小鼠在这三种状态间转换时,腺苷浓度的变化相对较快。胞外腺苷浓度的快速变化揭示其释放可能依赖于神经活动(图2)。

先前已有研究表明,BF胆碱能神经元(ChAT+)和谷氨酸能神经元(VGLUT2+)在觉醒和REMS阶段非常活跃,而且利用光遗传学技术激活这两类神经元可以促进机体觉醒。因此,作者首先通过检测ChAT+神经元活动与胞外腺苷浓度变化的相关性研究了BF ChAT+神经元的作用。作者利用“两侧双探针”法将AAV2/9-hsyn-GRABAdo1.0和AAV2/9-EF1α-DIO-GCaMP6s分别注入ChAT-Cre小鼠大脑半球两侧BF,2周后测定荧光信号,实验数据表明,ChAT+神经元钙活动与胞外腺苷浓度的变化高度相关,且钙活动通常早于腺苷信号变化~24 s,表明BF ChAT+神经元可能调控胞外腺苷的浓度变化。
接着,作者将AAV2/9-hsyn-GRABAdo1.0和AAV2/9-hSyn-FLEX-ChrimsonR-tdTomato 混合注射ChAT-Cre小鼠BF,光遗传激活ChAT+神经元,利用光纤记录腺苷的释放量。ChAT+神经元的激活引起胞外腺苷小幅显著增加,表明ChAT+神经元可调节睡眠觉醒周期中胞外腺苷浓度的变化。但是,诱发腺苷浓度变化的幅度在不同试验中有较大差异,表明BF中可能存在其它类型神经细胞调节睡眠觉醒周期中胞外腺苷浓度的变化(图3)。
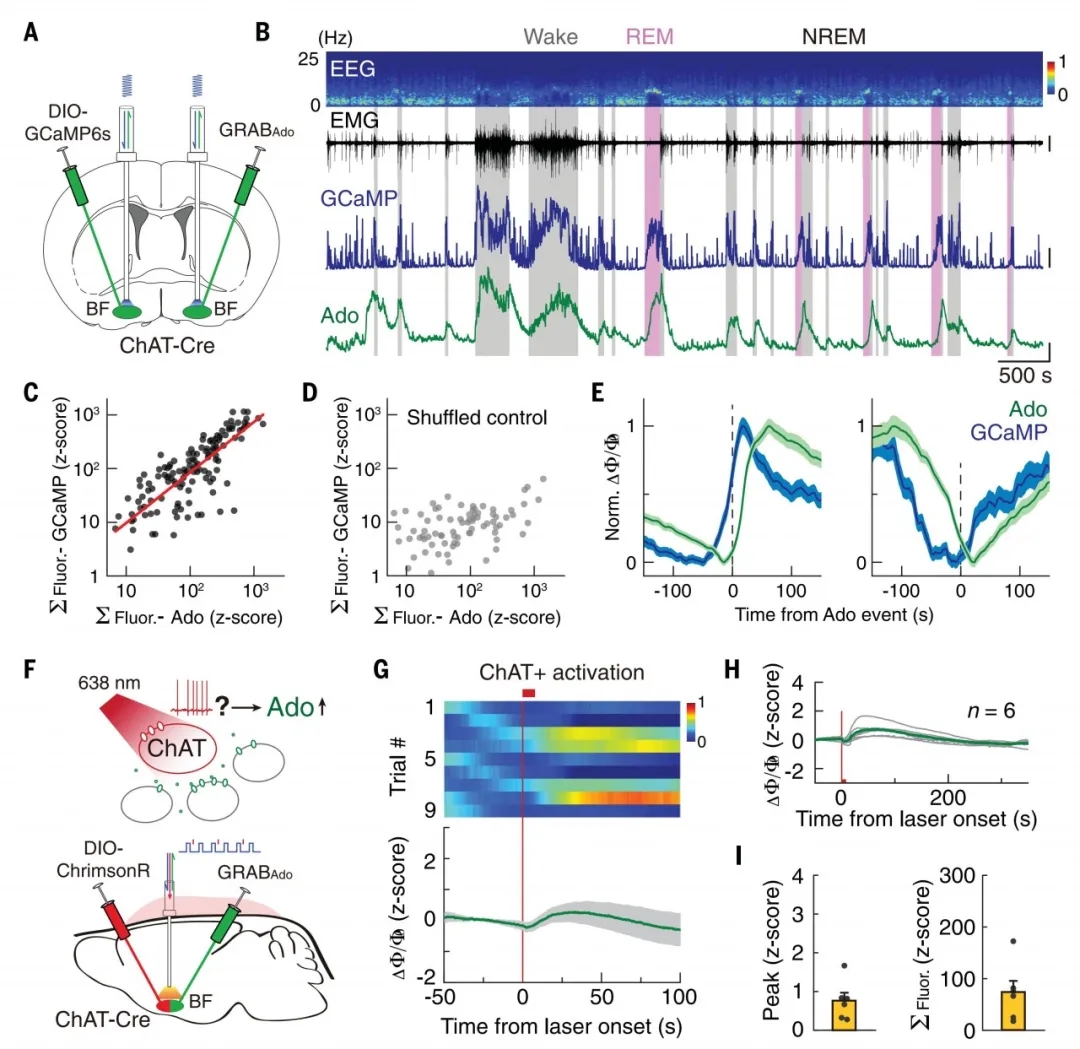
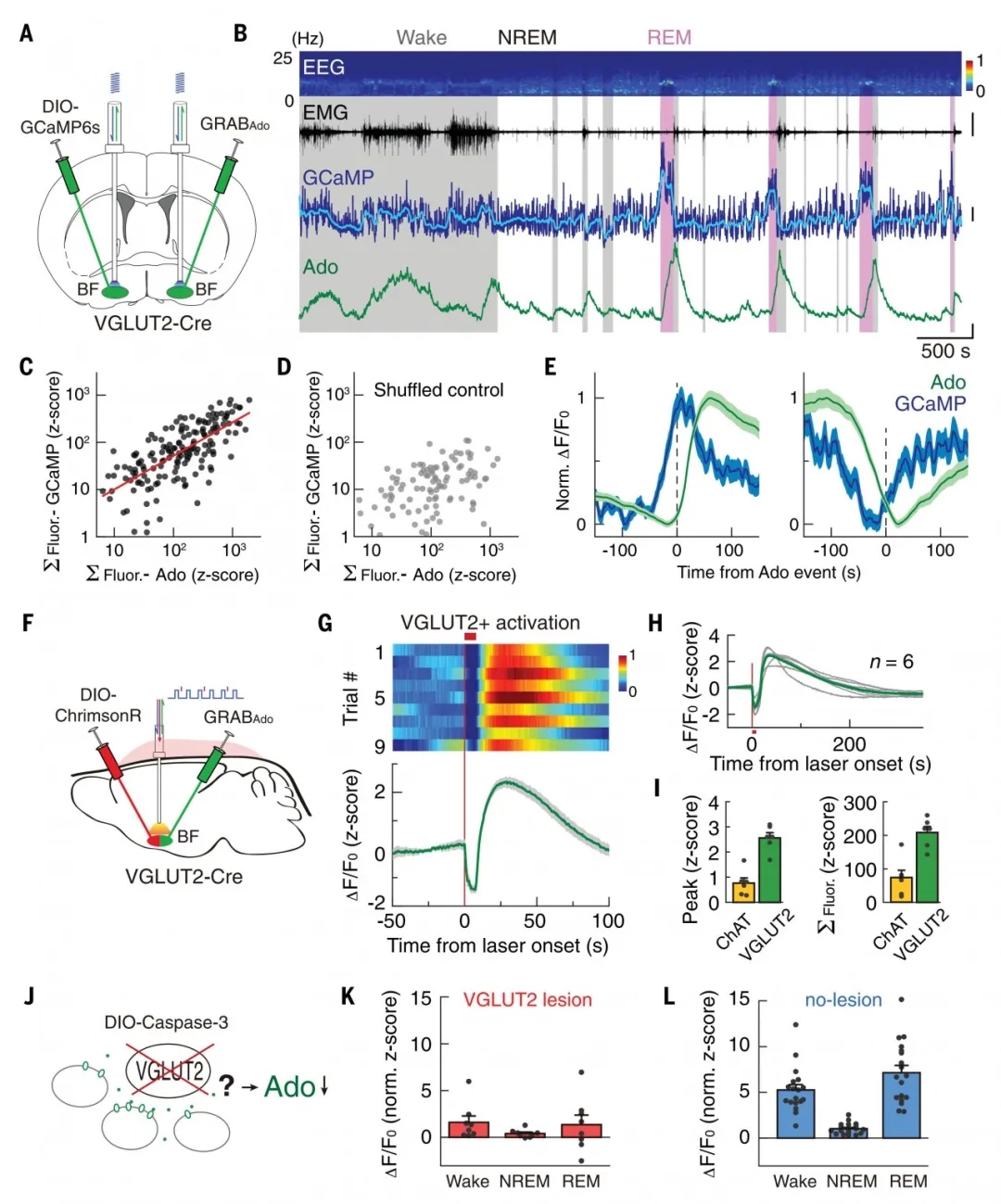
随后,作者采用同样的“两侧双探针”法又进一步研究了BF VGLUT2+神经元的调节作用。实验数据表明,BF VGLUT2+神经元中的钙活动与胞外腺苷浓度变化高度相关,且早于腺苷信号~41s。光遗传激活VGLUT2+神经元可引起胞外腺苷浓度大幅增加,并且高于激活ChAT+神经元所引起的胞外腺苷浓度,这表明VGLUT2+神经元对调节睡眠觉醒周期中胞外腺苷浓度的变化贡献更大。
接下来,作者用Caspase-3特异损毁BF VGLUT2+神经元,测量腺苷浓度的变化。将AAV2/9-hsyn-GRABAdo1.0和AAVDJ-flex-taCasp3-TEVp混合注射VGLUT2-Cre小鼠BF,两周后观察到BF VGLUT2+神经元数量显著减少。与对照组相比,实验组小鼠BF胞外腺苷浓度的增加在觉醒和REM阶段显著减少。上述结果进一步证明BF VGLUT2+神经元对胞外腺苷浓度增加起重要调节作用(图4)。
最后,作者通过细胞特异性损毁验证了BF中VGLUT2+神经元的缺失会影响小鼠睡眠稳态。与对照组相比,BF VGLUT2+神经元缺失的小鼠在觉醒状态下的时间明显更长,且这种差异主要表现为小鼠在夜晚清醒度增加。
作者又检测了BF VGLUT2+神经元的缺失是否会影响睡眠恢复。亮灯6h使老鼠保持清醒状态,随后18h内进行恢复性睡眠。实验组小鼠NREM睡眠明显减少,并且NREM睡眠时间下降速度明显加快。上述结果表明BF VGLUT2+神经元的缺失影响了小鼠的睡眠稳态(图5)。

此研究中,作者开发了一种新型的高灵敏度、高特异性和高时间分辨率的腺苷探针(GRABAdo),通过结合光遗传学操作和细胞特异损毁技术,阐明了睡眠觉醒周期中小鼠基底前脑谷氨酸能神经元调节胞外腺苷浓度变化,积累睡眠压力,从而对睡眠稳态具有重要调节作用。该研究为机体在清醒状态下神经活动通过刺激促眠因子的释放而增加睡眠压力的机制提供了新的见解。
原文链接:https://science.sciencemag.org/content/369/6508/eabb0556
| AAV病毒 | AAV2/9-hsyn-GRABAdo1.0-mScarlet; AAV2/9-hsyn-GRABAdo1.0 ; AAV2/9-hsyn-GRABAdo-mut; |
| 注射动物 | 7周龄以上雄性和雌性小鼠 |
| 注射体积 | 0.2 ~ 0.4 μl |
| 注射方式 | 脑立体定位注射 |
| 注射位置 | 基底前脑 |
| 检测时间 | 2周 |
| 探针类别 | 病毒名称 |
|---|---|
| 胆碱类 | 乙酰胆碱探针 |
|
AAV9-hSyn-GACh2.0 |
|
|
AAV9-TRE-GACh2.0 |
|
| AAV9-hSyn-ACh3.0(ACh4.3) | |
| AAV9-CaMKII-ACh3.0(ACh4.3) | |
| AAV9-GfaABC1D-ACh3.0(ACh4.3) | |
| …… | |
| 嘌呤类 | 腺苷探针 |
|
AAV9-hsyn-Ado1.0(B10) |
|
| AAV DJ-hsyn-Ado1.0(B10) | |
| AAV9-EFS-DIO-Ado1.0 | |
| AAV9-GfaABC1D-Ado1.0 | |
|
AAV9-hsyn-Ado1.0mut(B10-F168A) |
|
| …… | |
| 腺苷三磷酸探针 | |
| AAV9-hsyn-ATP1.0(B09) | |
| AAV DJ-hsyn-ATP1.0 | |
| AAV9-EFS-DIO-ATP1.0 | |
| AAV9-GfaABC1D-ATP1.0 | |
| AAV9-hsyn-ATP1.0mut(B09 N283A) | |
| …… | |
| 生物胺类 | 多巴胺探针 |
|
AAV9-hSyn-DA1h(DA4.2) |
|
|
AAV9-hSyn-DIO-DA1h(DA4.2) |
|
|
AAV9-hSyn-DA1hmut(DA4.2mut) |
|
| AAV9-TRE-DA1hmut(DA4.2mut) | |
| AAV9-CaMKII-DA1h(DA4.2) | |
| …… | |
| 五羟色胺探针 | |
| AAV9-hsyn-r5HT1.0 | |
| AAV9-CAG-r5HT1.0 | |
| AAV9-hsyn-r5HT2.0 | |
| AAV9-hsyn-5HT2.1(renamed as 5-HT1.0) | |
| AAV9-hsyn-DIO-5HT2.1(renamed as 5-HT1.0) | |
| …… | |
| 组胺探针 | |
| AAV9-hsyn-HA1.0h (His1.0) | |
| AAV9-hsyn-HA1.0mut (His1.0mut) | |
| AAV9-hsyn-HA1.0m (His1.0M) | |
| AAV9-hsyn-DIO-HA1.0h | |
| AAV9-hsyn-HA1.0h-mScarlet | |
| …… | |
| 去甲肾上腺素探针 | |
|
AAV9-hsyn-NE1m(NE2.1) |
|
|
AAV9-hSyn-DIO-NE1m(NE2.1) |
|
|
AAV9-hSyn-NEmut(NE2.1mut) |
|
|
AAV9-TRE-NE1m(NE2.1) |
|
| AAV9-hsyn-mRuby3-NE1m(NE2.1) | |
| …… | |
| 神经肽类 | 血管活性肠肽探针 |
| AAV9-hsyn-VIP1.0 | |
| AAV9-hsyn-DIO-VIP1.0 | |
| …… | |
| 神经肽Y探针 | |
| AAV9-hsyn-NPY1.0 | |
| AAV9-EF1a-DIO-NPY1.0 | |
| …… | |
| 神经降压肽探针 | |
| AAV9-hsyn-NTS1.0 | |
| …… | |
| 胆囊收缩素 | |
| AAV9-hsyn-CCK2.0 | |
| AAV1-hsyn-CCK2.0 | |
| AAV9-hsyn-DIO-CCK2.0 | |
| …… | |
| 促肾上腺皮质激素释放激素探针 | |
| AAV9-hSyn-CRF3.0 | |
| AAV9-EFS-DIO-CRF3.0 | |
| AAV9-CaMKII-CRF3.0 | |
| AAV9-hSyn-CRFmut | |
| AAV9-EFS-DIO-CRFmut | |
| …… | |
| 精氨酸血管加压素探针 | |
| AAV9-hsyn-AVP2.0 | |
| AAV9-hsyn-DIO-AVP2.0 | |
| …… | |
| 催产素探针 | |
| AAV9-hSyn-OXT1.0 | |
| AAV9-EFS-DIO-OXT1.0 | |
| AAV9-hSyn-OXTmut | |
| AAV9-EFS-DIO-OXTmut | |
| …… | |
| 生长激素抑制素探针 | |
| AAV9-hsyn-SST2.0 | |
| AAV9-hsyn-DIO-SST2.0 | |
| …… | |
| 其他 | 大麻素AEA探针 |
| AAV9-hsyn-AEA1.0 | |
| …… | |
| 内源性大麻素eCB探针 | |
| AAV9-hsyn-eCB2.0 | |
| AAV9-hsyn-DIO-eCB2.0 | |
| AAV9-hsyn-eCB2.0mut | |
| AAV9-hsyn-DIO-eCB2.0mut | |
| AAV9-GfaABC1D-eCB2.0 | |
| …… | |
| 褪黑素探针 | |
| AAV9-hsyn-MT2.0 | |
| …… | |
| 以上探针均由维真生物协助提供,其中已发表的探针(蓝色标注),点进进入了解更多探针详情。欢迎大家点击“AAV工具” 中的“神经递质探针”进行选购!!如若您对探针序列或者未发表的探针感兴趣,请您联系北京大学李毓龙课题组(yulonglilab2018@gmail.com)。 | |
维真生物还可为您提供 光遗传学、化学遗传学、钙指示剂、Cre/Flp重组酶 等工具AAV,欲知更多相关产品请拨打技术热线电话400-077-2566。

400-077-2566

service@wzbio.cn







